- 移动端


北京索莱宝科技有限公司品牌商
14 年
手机商铺
- NaN
- 0
- 0
- 2
- 2

Solarbio
试剂
已认证品牌介绍

Oxoid
试剂
已认证品牌介绍

greiner bio-one
培养
已认证品牌介绍
Solarbio
试剂
已认证品牌介绍
Oxoid
试剂
已认证品牌介绍
greiner bio-one
培养
已认证品牌介绍
Solarbio
试剂
已认证品牌介绍
Oxoid
试剂
已认证品牌介绍
greiner bio-one
培养
已认证品牌介绍
推荐产品



公司新闻/正文
文献解读|植物和动物正单链 RNA 病毒在其负义链上编码对病毒侵染重要的小蛋白
2693 人阅读发布时间:2024-02-06 14:00
大多数感染真核生物的病毒都有一个正单链 RNA(+ssRNA) 基因组。一般认为,+ssRNA 病毒只在基因组正链 RNA(+RNA) 中编码开放阅读框架 (ORF),而病毒的负义链 RNA(-RNA) 是一种病毒复制中间体,其功能是作为后代基因组 RNA 生物合成的模板。病毒基因组 RNA 作为 mRNA 直接与核糖体相互作用,并被翻译成病毒蛋白质,包括用于病毒复制的 RNA 依赖的 RNA 聚合酶 (RdRPs)。
植物病毒具有密集的基因组,小开放阅读框 (sORFs) 是编码长度小于 100 个氨基酸 (aa) 蛋白质的开放阅读框;sORFs 翻译的小蛋白与不同生物体(包括植物和动物病毒)的各种生物过程有关。马铃薯 Y 病毒科目前由 12 个属、237 个物种组成,其中包括最大的植物感染 +ssRNA 病毒(马铃薯 Y 病毒)。约 10kb 的马铃薯病毒基因组由编码大的多聚蛋白的单个 ORF 组成,该蛋白被蛋白水解加工成 10 种功能蛋白,分别为 P1、HC-Pro、P3、6K1、CI、6K2、VPg、NIa-Pro、NIb 和衣壳蛋白质 (CP)。另一种蛋白质 P3N-PIPO 是通过转录滑移合成的,这导致在一小部分 RNA 转录物的 5’ 端附近增加了额外的 A 残基。但马铃薯 Y 病毒属是否在其 RNA 中编码额外的小蛋白尚未被探索。
冠状病毒(冠状病毒科)是一个有包膜 +ssRNA 病毒的大家族。严重急性呼吸综合征冠状病毒 2 型 (SARS-CoV-2) 属于冠状病毒科β冠状病毒属。SARS-CoV-2 的基因组大小约为 30kb,包含 14 个典型的病毒开放阅读框,50 个 ORF1a/ORF1ab 组成了整个基因组的前三分之二,它编码一种多聚蛋白,最终被蛋白酶切割成 16 种非结构蛋白 (Nsp1-Nsp16),病毒基因组 3’ 端的最后三分之一包含 13 个 ORF,编码 4 种结构蛋白(刺状蛋白 [S]、包膜蛋白 [E]、膜蛋白 [M] 和核衣壳蛋白 [N])和 9 种可能的辅助蛋白 (ORF3a、ORF3b、ORF6、ORF7a、ORF7b、ORF8、ORF9b、ORF9c 和 ORF10)。但 SARS-CoV-2 的 RNA 中是否编码额外的小蛋白仍不清楚。
本文在来自不同科的几种植物、脊椎动物和无脊椎动物 +ssRNA 病毒的 RNA 中鉴定了几个潜在的小反向 ORFs(rORFs)。通过研究芜菁花叶病毒 (TuMV),发现这些 sORFs 对病毒侵染至关重要,其编码的蛋白质具有特定的亚细胞定位,证明了 rORF2 蛋白是由马铃薯 Y 病毒 rORFs 编码的蛋白,可与病毒复制复合体共定位以及 TuMV RdRP 相互作用,并作为毒力因子发挥作用。动物病毒 SARS-CoV-2 的 rORFs(rORF1-rORF3) 可能通过拮抗 I 型干扰素 (IFN-I) 介导的抗病毒先天免疫反应来促进水泡性口炎病毒 (VSV) 感染。另外,RNA 中的 TuMV-rORF 可能是由病毒内部核糖体进入位点 (IRES) 翻译的。因此,作者提出 +ssRNA 病毒在其-RNA 中编码小功能蛋白,这些蛋白可以影响病毒复制和毒力,或者可能有助于逃避宿主先天免疫反应,从而促进病毒侵染。这些发现扩展了对 +ssRNA 病毒所采用的编码策略的理解,并扩大了已知的病毒蛋白质组及其与宿主细胞的潜在相互作用。
基本信息
题目:
Plant and Animal Positive-sense Single-stranded RNA Viruses Encode Small Proteins Important for Viral Infection in Their Negative-sense Strand
期刊:Molecular Plant
影响因子:27.5
PMID: 37777826
DOI: 10.1016/j.molp.2023.09.020
通讯作者:周雪平 李方方
作者单位:中国农业科学院植物保护研究所
索莱宝合作产品:
|
产品货号 |
产品名称 |
|
K200015M |
Anti-mCherry Tag Monoclonal Antibody |
摘要
正单链 RNA(+ssRNA) 病毒是自然界中含量最丰富的真核生物病毒,需要使用基因组 +RNA 作为复制模板合成-RNA。植物和动物 +ssRNA 病毒的 RNA 中含有小的开放阅读框 (ORF)(反向 ORF[rORF])。利用芜菁花叶病毒 (Tumv) 作为植物 +ssRNA 病毒的模型,证明了 rORFs 编码的小蛋白具有特定的亚细胞定位,并通过质谱分析证实 rORF2 在感染细胞中的存在。TuMV rORF2 编码的蛋白质形成点状颗粒,定位于核周区域,并与病毒复制复合体共定位。RORF2 蛋白可以直接与病毒 RNA 依赖的 RNA 聚合酶相互作用,rORF2 的突变完全消除了病毒的侵染,而 rORF2 的异位表达拯救了突变的病毒。此外,SARS-CoV-2 的 RNA 具有抑制 I 型干扰素产生和促进水泡性口炎病毒侵染的作用。作者提供了 TuMV 可能利用内部核糖体进入位点来翻译这些小 rORF 的证据。综上所述,这些发现表明 +ssRNA 病毒的-RNA 也可以具有编码能力,其中编码的小蛋白在病毒侵染中发挥关键作用。
研究内容及结果
1.鉴定 +ssRNA 病毒 RNA 中的 sORFs
马铃薯 Y 病毒科、南方菜豆花叶病毒科、冠状病毒科和黄病毒科病毒的基因组序列分析发现了许多编码超过 10 个氨基酸的蛋白质的 ORFs,与 +RNA 中已知的 ORFs 类似,这些病毒的 RNA 在每个科内选定的病毒子集中具有高度代表性(图 1A–D)。由于这些 ORFs 是在病毒 +RNA 的反向互补序列中预测的,所以将它们命名为 rORFs(图 1E)。
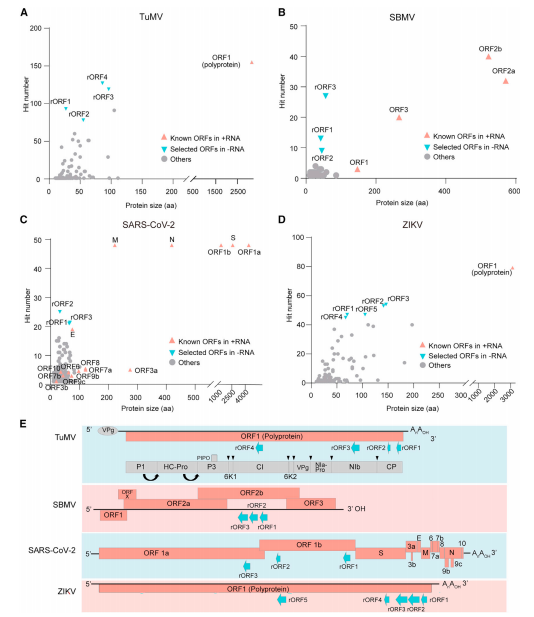
图 1
2.TuMVrORF 编码的蛋白质具有特定的亚细胞定位,可被招募到病毒复制复合体(VRC)
在 TuMV 中鉴定出几个小 rORFs、rORF1-rORF4,并发现在马铃薯 Y 病毒科中有大量匹配(图 1A 和 2A)。TuMV-rORF1–rORF3 编码的蛋白质与黄色荧光蛋白 (YFP) 融合,定位于细胞核、核周区域和细胞质,而 rORF4 仅限于核周区域(图 2B)。TuMV rORF1-YFP 定位在叶绿体周围,并与过氧化物酶体标记 DsRed-SKL 共定位,但不与高尔基体标记共定位(图 2C),与叶绿素自发荧光和 ER 标记 mCherry-HDEL(his-asp-glu-leu 在其 N 末端融合 mCherry 蛋白)的共定位发现 TuMVrORF3-YFP 在叶绿体和内质网 (ER) 中(图 2D)。马铃薯 Y 病毒科复制发生在细胞质膜结合的 VRC 中,在细胞核附近可以看到密集的块状物。使用表达青色荧光蛋白 (CFP) 标记的病毒 RdRP(NIb) 和 mCherry 标记的叶绿体定位膜蛋白 (6K2) 的 TuMV-6K2-mCherry-CFP-NIb 感染性克隆(图 2E),可以发现这些 rORF 编码的蛋白质在病毒侵染过程中被招募到 VRC 中复制(图 2F)。四种新蛋白均与 6K2 诱导的 TuMVVRC 和 dsRNA 共定位,这些 rORF 在病毒复制过程中 VRC 的形成和活性中具有潜在作用(图 2G-H)。
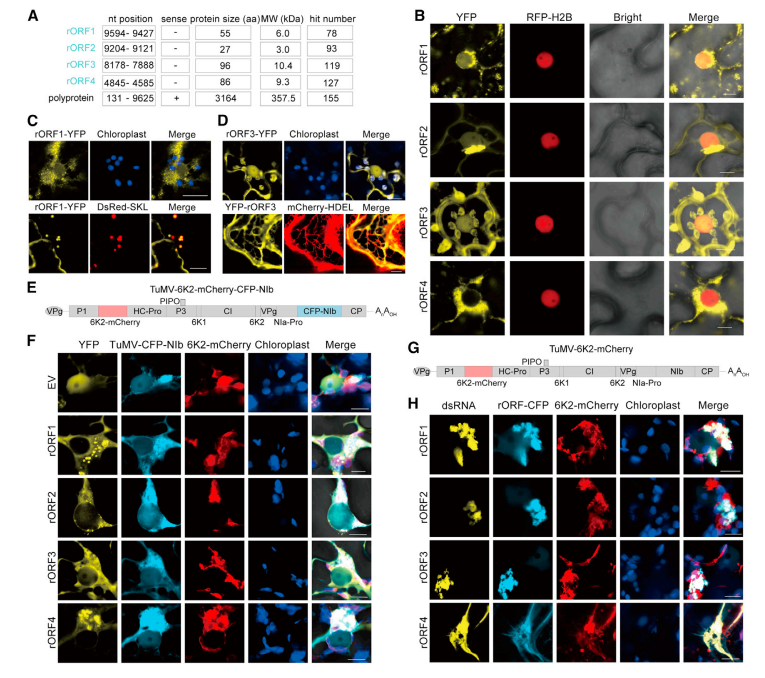
图 2
3.TuMV rORF2 编码蛋白是病毒感染所需的毒性因子
所有突变病毒在接种后 60h 时均呈现出明显较少的 RNA 和蛋白质积累(图 3A-C)。接种后 5d 和 12d,TuMV-GFP-mrORF2 未能在接种植物的全身组织中检测到病毒 RNA 和 CP;rORF1、rORF3 或 rORF4 的突变对 TuMV 症状和病毒积累的影响较轻微(图 3D-H)。
除 P3NPIPO 外,rORF2 蛋白与已报道的 TuMV 蛋白同时被检测到(图 4A-B)。PVX-rORF2 在 28d 时表现出严重的症状(图 4C)。此外,PVX 载体表达的 rORF2 基因可以弥补接种 TuMV-GFPmrORF2 突变体的本氏烟草中 rORF2 的缺失(图 4D-F)。RORF2-YFP 的转基因表达恢复了 TuMV-mrORF2-GFP 的感染(图 4G-I)。
rORF2 和 11 种典型 TuMV 蛋白酵母双杂交 (Y2H) 测定发现 rORF2 编码的蛋白质与酵母细胞中的 NIb 相互作用(图 4J)。红色荧光蛋白 (RFP)-H2B 转基因本氏烟草植物叶子的双分子荧光互补 (BiFC),证实体内相互作用发生在植物细胞的细胞质中(图 4K)。NIb 与 TuMV 感染细胞 VRC 中的 rORF2 相互作用(图 2F 和 4K),进一步表明 rORF2 参与 TuMV 复制。
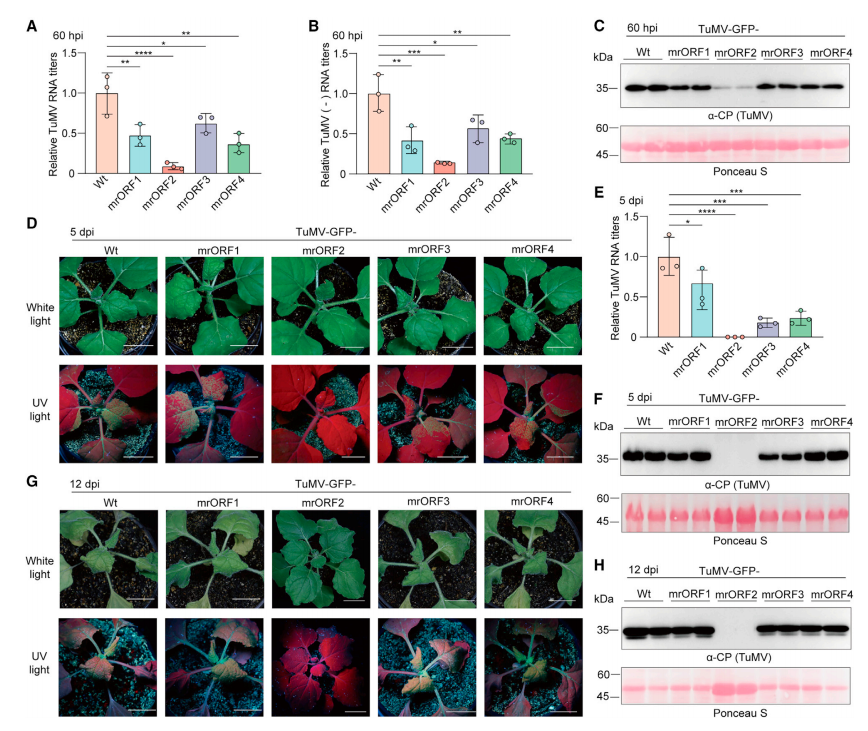
图 3
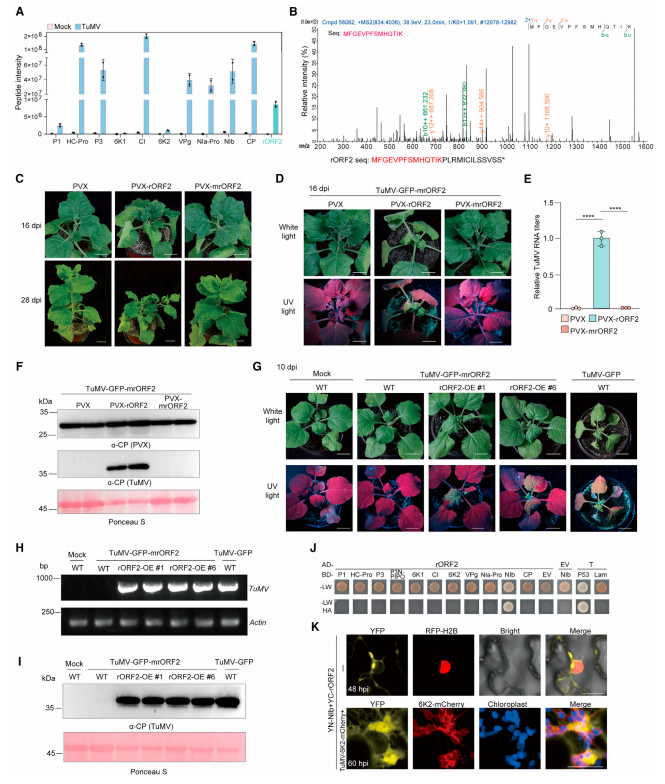
图 4
4.SARS-CoV-2rORF 的特征
使用 SARS-CoV-2 作为模型(图 5A)推测由 rORFs 编码的蛋白质尺寸较小 (<10kDa),rORFs 在 SARS-CoV-2 相关变体 (VOC) 中也完全保守(图 5B)。rORF3 编码的蛋白质预计包含两个跨膜结构域(图 5C)。SARS-CoV-2 rORF 编码蛋白的 GFP 标记,三种融合 GFP 的 rORF 编码蛋白均积累到相当的水平,但在 HEK293T 细胞中出现不同的亚细胞定位。GFP-rORF1 分布在细胞核和细胞质中,GFP-rORF2 主要位于细胞核中,GFPrORF3 主要分布在细胞质中(图 5D-E)。
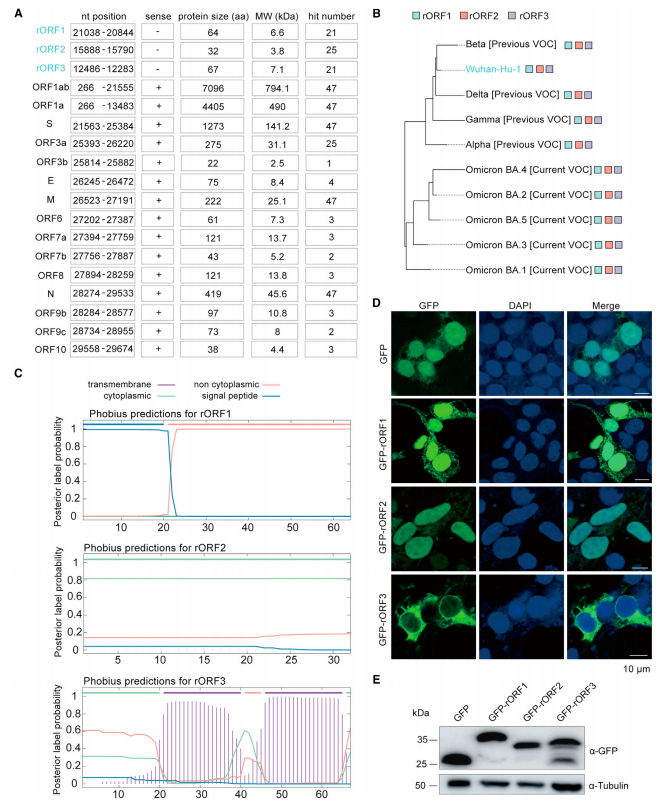
图 5
5.SARS-CoV-2rORF 编码蛋白是 IFN-I 拮抗剂
标记 SARS-CoV-2 的 rORF1-3 可以降低维甲酸诱导基因 IRIG-IN 诱导的干扰素β启动子活性(图 6A)。SARS-CoV-2 rORF2 和 ORF6 可明显抑制外源干扰素-α 诱导的 ISRE 启动子的活性(图 6B)。
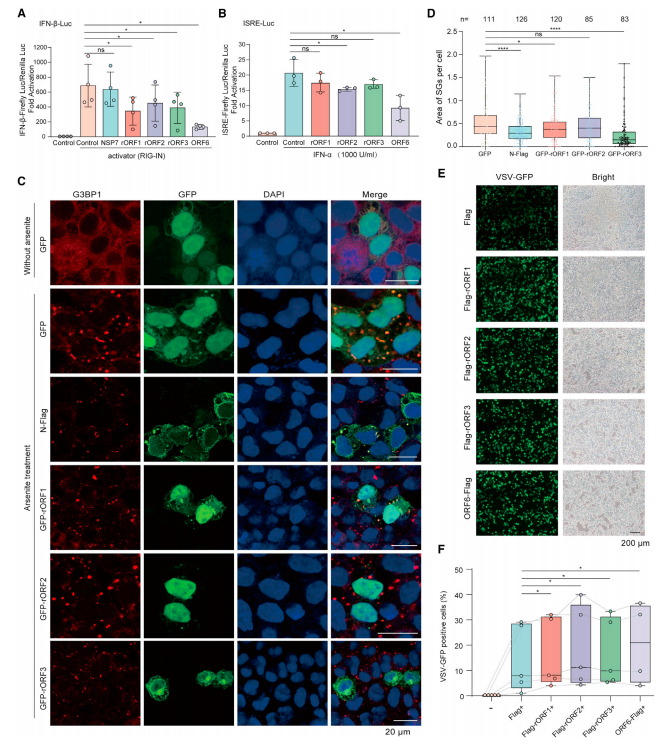
图 6
6.SARS-CoV-2rORF 编码蛋白抑制 SG 的形成并促进病毒侵染
与表达 GFP 的细胞相比,表达 N-FLAG 的细胞中每个细胞可检测的 SG 面积显著减少,并且 N 蛋白与 SG 中的 G3BP1 共定位,表达 rORF1-GFP 和 rORF3-GFP 的细胞中 SG 形成显著减少(图 6C 和 6D),表明 rORF1 和 rORF3 介导对 SG 形成的抑制。GFP-rORF1 和 G3BP1 也共定位于 NaAsO2 处理细胞中的剩余 SG(图 6C),表明 rORF1 可能通过与 G3BP1 相互作用隔离 G3BP1 来减弱 SG 形成。表达单个 SARS-CoV-2rORF 的细胞中 VSV-GFP 侵染的效率显著高于对照细胞(图 6E-F)。
7.小 TuMV rORF 可能由病毒 IRES 翻译
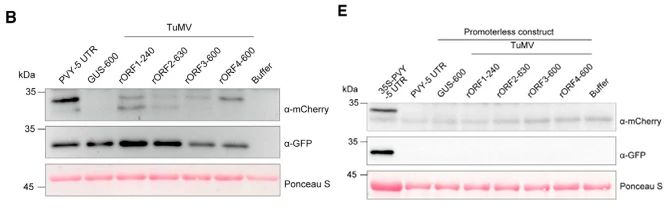
作者设计了一个双顺反子系统(图 7A)。通过农杆菌介导法将含有 TuMV rORFs 上游序列的双顺反子载体瞬时地表达到本氏烟草的叶片组织中(图 7B)。mCherry 积累的差异不是由于 mRNA 水平的变化所致(图 7C)。用β-葡萄糖醛酸酶 (GUS) 的序列取代驱动 GFP-mCherry 转录的 35S 启动子(图 7D)。蛋白质印迹分析表明无启动子载体无法在本氏烟草叶子中表达 GFP 或 mCherry(图 7E)。qRT-PCR 分析表明 35S 启动子驱动的表达 mRNA 产物在植物细胞中具有特异性条带,表明不存在隐性剪接位点(图 7F)。
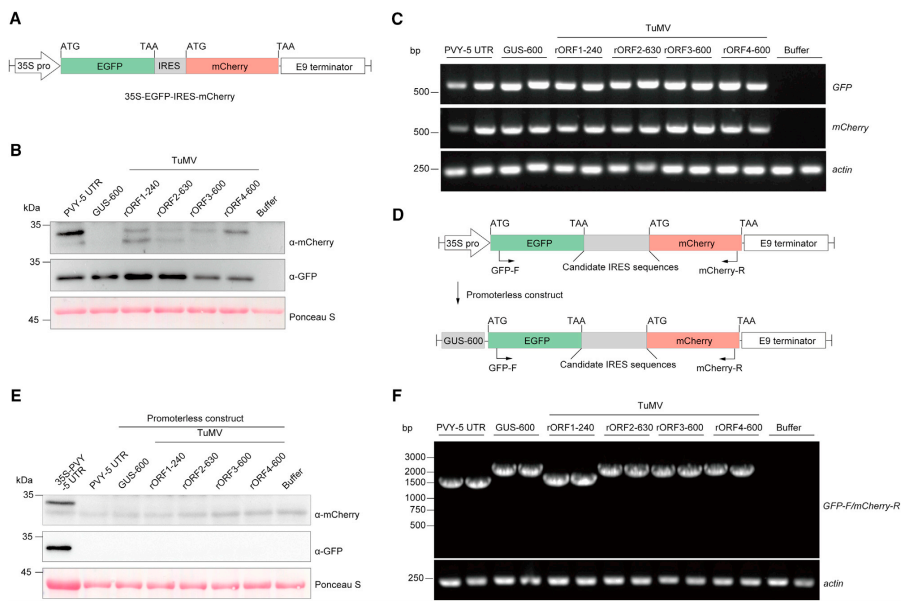
图 7
结论使用两种不相关的 +ssRNA 病毒感染不同来源的生物体,实验证明了 +ssRNA 病毒的 RNA 在病毒侵染过程中编码具有生物学作用的小蛋白。该研究需重新考虑之前的假设,即该链缺乏生物相关的蛋白质编码信息,如植物 TuMV 所示,需要重新研究已知的 +ssRNA-vi 病毒的蛋白质组,并且可能会扩大。
索莱宝产品亮点